Miura H. Mechanism underlying the in vitro formation of urine dysmorphic erythrocytes. Lab Med Int 2024 3(1): 2-7. doi: 10.51041/lmi.3.1_2
Review
Lab Med Int 2024 3(1): 2-7
† Department of Clinical Laboratory, Faculty of Medicine, Yamagata University, 2-2-2 Iida-Nishi, Yamagata-shi, Yamagata 990-2331, Japan.
E-mail: hmiura27″@”hotmail.com
Department of Clinical Laboratory, Faculty of Medicine, Yamagata University.
Received November 13, 2019; accepted March 1, 2022
Key Words
dysmorphic erythropoiesis mechanism, glomerular bleeding, urinary acanthocytes, urinary osmolality, hematuria
I. Introduction
In 1979, Birch et al.1) reported that hematuria can be broadly divided into glomerular (dysmorphic erythrocytes) and urinary tract-derived (isomorphic erythrocytes) based on the morphology of erythrocytes. This classification is also widely used in diagnostics and laboratory tests. On the other hand, various alterations have been reported in the mechanism of erythropoiesis, but no consensus has been reached. There are three of his hypotheses about the formation mechanism of dysmorphic erythrocytes: (1) Mechanical hypothesis: Caused by leakage from damaged glomerular basement membrane, (2) Nephron passage hypothesis: Caused by sudden changes in osmotic pressure, pH, and urinary components in nephron, (3) Mixed hypothesis: First A combination of his two hypotheses. To test these hypotheses, the author searched his original papers and reviews on the mechanism of dysmorphic erythrocyte formation published in his PubMed from 1979 to 2018.
II. Result
1. Documents on which these hypotheses are based
In references 2)3), dysmorphic erythrocytes leaking from the damaged glomerular basement membrane are observed, but this is not a report on the formation mechanism of dysmorphic erythrocytes. It proves that the cause of hematuria bleeding is the injured glomerulus.
Stapleton 4) reports that mechanical trauma may explain the dysmorphic red blood cells that represent glomerulonephritis. Later, Schramek 5) introduced a hypothesis about the formation mechanism of dysmorphic erythrocytes.
2. About three hypotheses
Hypothesis (1)
Kubota 6) reported that he could produce donut-shaped dysmorphic erythrocytes simply by filtering erythrocytes soaked in saline with a 5 μm fibrin or an untreated 3 μm
filter (38℃, 30 minutes). However, the full picture of dysmorphic red blood cells and acanthocytes (ACs) is not displayed.
Daza JL 7) showed an image (Masson’s trichrome staining) of erythrocytes passing through the glomerular basement membrane. Daza JL also explained that the causes of erythrocyte malformations are mechanical trauma as cells pass through cracks in the glomerular basement membrane and osmotic trauma as cells pass through nephrons. This indicates that hypothesis (3) is taken into account as the cause of dysmorphic erythropoiesis.
In 2015, Yuste C et al. 8) “Review: Pathogenesis of glomerular hematuria” stated as follows. The presence of dysmorphic red blood cells with irregular contours and shapes in the urine is a characteristic symptom of glomerular hematuria. It also suggests deviations from glomerular capillaries to the urinary tract. Therefore, glomerular hematuria is a marker of glomerular filtration rate barrier (GFB) dysfunction or damage. This indicates that hypothesis (1) is considered to be the formation mechanism of dysmorphic erythrocytes.
Hypothesis (2)
If hypothesis (2) is defined as the case where erythrocytes are continuously immersed in the osmotic pressure of simulated urine or nephron solution, Schramek 2), Miura et al 9) 10) 11), Kitamoto et al. 12) We carried out a nephron simulation. The authors of this method, Miura and Kitamoto, were able to produce dysmorphic red blood cells in patients with glomerulonephritis. Schramek, on the other hand, said that it is not possible to produce dysmorphic erythrocytes with this method, and that after this method, additional hemolytic environments can produce dysmorphic erythrocytes.
Rath 13) soaked erythrocytes in solutions with different osmotic pressure and pH, but no dysmorphic erythrocytes were produced. In response to this result, Rath reported that hypothesis (3) was necessary. When red blood cells are immersed in a single urine or solution, only individual osmotic forms are obtained. Therefore, this does not seem to prove hypothesis (2).
Hypothesis (3)
Briner 14), urine (pH 5-8, osmolality 200-800 mOsm) and blood mixture were aspirated through a 3 μm polycarbonate filter and then incubated at 37℃. for 1 hour. Of these mixtures, Figure 1 presented by Briner was obtained after aspiration under conditions of resuspension in urine with a pH of 7.0 and an osmotic pressure of 300 mOsm. Also, no dysmorphic red blood cells were produced without filtration. In addition, the debate states that the formation of dysmorphic red blood cells requires two factors: the passage of cracks and the passage of certain suspensions.
From their results, it is unlikely that hypothesis (3) can explain the formation mechanism of modified erythrocytes. It may be the effect of 1 hour of incubation or fixation. Furthermore, glomerular red cells shown by Briner (14) are the same as the “ACs-like cells” experienced by the authors, and care must be taken when classifying the morphology, and it is important to recognize them as isomorphic erythrocytes. This means hypothesis (3), which is a mixture of the mechanical hypothesis and the nephron transit hypothesis, and it is believed that only one solution (pH 7.0, osmotic pressure 300 mOsm) produces dysmorphic erythrocytes. not. There are also ACs, ring cells, and target cells, and from the fact that the overall picture of the morphology is shown, it is considered to be a true image of dysmorphic red blood cells. The author thinks that this image is the dysmorphic red blood cell itself, which is the same as the proof of hypothesis (3).
3. Papers other than the three hypotheses
Halbhuber 15) states that treatment with proteases prior to the nephron simulation increased the number of dysmorphic erythrocytes from 35% to 80% compared to untreated cells (12% -15%). However, the fact that 12% to 15% of dysmorphic erythrocytes were produced without protease treatment proves hypothesis (2). However, Miura and Kitamoto have succeeded in forming dysmorphic erythrocytes without pretreatment with proteases.
Proteases degrade membrane skeletal proteins (ankyrin, band 4.1 protein), band 3 proteins, and glycophorin C, interfering with the interaction of spectrins with membranes and other membrane scaffold proteins. This suggests that as the degradation progresses, the bond between the erythrocyte membrane and the spectrin loosens, and the erythrocyte membrane structure loosens. It is thought that the loosening of these erythrocyte membrane structures results in the formation of protrusions at high osmotic pressure. Therefore, pretreatment with proteases may promote dysmorphic erythropoiesis.
On the other hand, since thrombin is a kind of proteolytic enzyme, thrombin may be involved. In a review entitled “Protease-activated Receptors (PAR) in the Progression of Renal Disease,” he reported by Palygin16) that thrombin is present in the urine of patients with glomerulonephritis rather than in the urine of healthy individuals. In addition, this study showed that excessive PAR stimulation can lead to excessive intracellular Ca2 + levels and cell apoptosis, followed by proteinuria and glomerular damage, which may be directly involved in the progression of renal disease. It was.
In addition, “glomerulonephritis is characterized by carbonyl stress and elevated methylglycoloxol (MGO), where erythrocyte suspensions and MGO accumulate Ca in the body.” Degrell et al. 17) We point out that factors may play an important role in the formation of dysmorphic erythrocytes in glomerulonephritis. These facts may fundamentally overturn the hypothesis about the formation mechanism of dysmorphic erythrocytes. However, without treatment with proteases or MGO, Miura et al. Produced dysmorphic erythrocytes in a solution containing only the urinary components Na, Cl, K, IP and urea (see below) and therefore in the urine of patients with glomerulonephritis. It is desirable to measure the content of protease (thrombin) and MGO in the body to clarify the relationship with dysmorphic erythropoiesis, and at the same time, it may be possible to make an early diagnosis of glomerulonephritis by evaluating these factors. Interesting.
Recently, Kitamoto et al. 18) reported that measurement of urinary thrombin enables early diagnosis of crescent glomerulonephritis (CresGN). According to this study, (1) in CresGN, fibrin deposition and thrombinuria were associated with extraglobulinal capillary tissue in which monocytes / macrophages express tissue factor. (2) Fibrin deposition in the CresGN glomerulus indicates thrombin formation. Therefore, the author hypothesized that thrombin was excreted in the urine and that certain he was a CresGN biomarker. (3) Thrombinuria was specific to glomerular inflammation and was not affected by systemic inflammation or coagulation. (4) It has been shown that thrombin produced at the glomerular bleeding site for hemostasis does not significantly affect the measurement of thrombinuria. ⑤ Thrombinuria showed high his CresGN specificity (90.1%) and moderate sensitivity (70.6%). The widespread use of early diagnostic measurement systems will be of great help in treating patients.
Therefore, in addition to the above three hypotheses, proteases or MGO may also be involved, as mentioned above. The mechanism of dysmorphic erythropoiesis remains unclear, justifying future research on this subject.
Finally, when experimenting with the three hypotheses, it is important to note the following:
*Hypothesis (1): Plasma → Bowman’s capsule → proximal tubule is isotonic pressure (285 mOsm). Therefore, the osmotic pressure of the immersion liquid before and after filtration should be isotonic urine or solution. We believe that the use of other osmotic pressures does not prove hypothesis (1).
*Hypothesis (2):
1; The osmotic pressure or pH of the urine or solution used is similar to the osmotic pressure of nephrons, but continuous immersion of red blood cells is essential.
2; The pH of hyperosmolar urine or solution used after a hemolytic environment must be acidic.
3; Hemolytic environmental conditions should be determined by the relationship between red blood cell count and osmotic pressure.
4; Morphological changes that occur after soaking in urine or solution require not only individual images, but also whole images containing dysmorphic red blood cells.
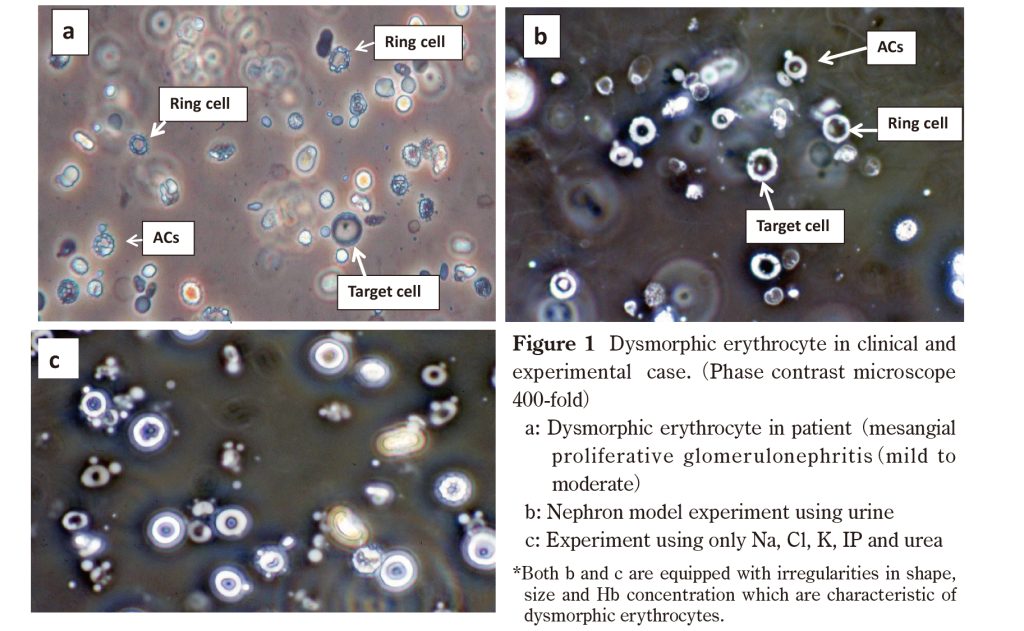
III. Discussion
As described above, various methods and interpretations have been attempted for the three hypotheses, and despite the fact that the electron micrographs and photomicroscopic images of erythrocytes deviating from the glomerular basement membrane have been shown as evidence of the cause of hematuria. However, it is undeniable that it has progressed to three hypotheses by being taken up as the hypothesis (1) of the cause of dysmorphic erythrocytes. Therefore, the factors that cause dysmorphic erythropoiesis are explained below.
1. About nephron osmotic pressure series
The osmotic pressure of each part of the nephron gradually changes in the following order. Bowman’s capsule and proximal tubule (285 mOsm), loop of Henle (1,200 mOsm), first distal tubule (50-100 mOsm: hemolytic environment), distal tubule-collecting duct (1,200 mOsm). Therefore, in this order, red blood cells need to be continuously (continuously) immersed in urine or solution.
2. About low osmotic pressure (hemolytic environment)
The osmotic pressure is 50 to 100 mOsm in the textbook, but experimentally it was 161 mOsm or less.
It was full. In addition, urea was needed to obtain different hemolysis levels (various Hb concentrations). Birch 19) states that changes in red blood cell morphology (the pattern of cell morphology is a polymorphism containing at least three different sizes or shapes) and hemoglobin (Hb) levels indicate glomerular hemorrhage. Of these, changes in Hb concentration cannot be explained by hypothesis (1). Regarding this hemolytic environment, Schramek et al. Suggested the existence of an endogenous causal factor released from hemolyzed erythrocytes, Briner et al. 14) found that hemolysis was closely related to the formation of deformation. Kitamoto et al. Report that hemolysis is not essential for the formation of dysmorphic erythrocytes, but the paper produces dysmorphic erythrocytes with only two types, low (140 mOsm) and hyperosmotic solution (1100 mOsm).
3. About urea
Kitamoto et al. Says that urea and other ion-containing solutes are thought to promote dysmorphic erythropoiesis. The authors found that urea caused varying Hb concentrations in dysmorphic erythrocytes, but NaCl did not. The difference in his Hb concentration in dysmorphic red blood cells is due to the difference in the amount of Hb released from hemolyzed red blood cells. It also depends on the degree of damage to the erythrocyte membrane. Dysmorphic erythrocytes produced by experiments using the author’s urine and solution met Birch’s requirements (Figure 1b, 1c).
4. About hyperosmotic pressure (dysmorphic erythrocyte formation site)
ACs (Figure 1a) are the most characteristic of the dysmorphic erythrocyte morphology found in the urine of patients with glomerulonephritis. In an in vitro nephron model experiment using urine, after immersing erythrocytes in his 95 mOsm (hemolytic environment), he developed ACs at a high osmotic pressure of 911 mOsm (Figure 1b). In experiments using only Na, Cl, K, IP, and urea, the 104 mOsm (hemolytic environment) solution was simply immersed in a 726 mOsm high osmotic solution (including K: 8.9, IP 9.3 mmol / L, respectively). Can generate ACs (Figure 1c). In this way, being able to make his ACs with only his five types of Na, Cl, K, IP, and urea made it easier to elucidate the causes of ACs. Therefore, ACs appeared in hyperosmotic urine or solution after passing through a hemolytic environment. It mimics the in vivo condition in which glomerular hemorrhage induces the formation of dysmorphic erythrocytes in the hyperosmolar region after the erythrocytes have passed through the hemolytic environment. Therefore, for dysmorphic erythropoiesis, it is essential to pass through the hemolytic environment and through the hyperosmolar region. Kitamoto et al. Also stated that conditions of pH 5 and osmotic pressure of 1,000 mOsm simulated in a collecting duct are essential for dysmorphic erythropoiesis. Therefore, it cannot be concluded that dysmorphic erythrocytes were produced simply from the experimental results of immersing erythrocytes in various mono osmotic urine or solutions. Hypothesis (2) is thought to be the result (image) obtained by the final hyper osmolarity in which erythrocytes are continuously immersed according to a nephron simulation. Hyper osmolarity required 425 mOsm or higher, and K or IP required 4.1 mmol / L and 4.3 mmol / L or higher, respectively. Also, no ACs were observed when the composition of the hyperosmolar solution was Na and Cl. However, the number of ACs increased with increasing osmolality or K and IP. These results suggest that the presence of K and IP promotes the formation of AC. However, the question arises as to whether K or IP is the main contributor to ACs formation. In this regard, the K + and PO43- concentrations in erythrocytes are as high as 140 mmol / L and 100 mmol / L, respectively. Some of these are thought to be eluted by hemolysis. For high osmotic solutions, ACs begin to appear at K 4.1 mmol / L and IP 4.3 mmol / L and above, producing large numbers of ACs at osmotic pressures 726 mOsm, K 8.9 mmol / L and IP 9.3 mmol / L and above, respectively. (Figure 1c). So far, KH2PO4 solution has been used in experiments, but the question is whether K + or PO43- is involved in the formation of ACs. Therefore, 87 mOsm low osmotic urine, KH2PO4 (pH 4.3), K2HPO4 (pH 8.3), KNO3 (pH 5.6), K2CO3 (pH 10.3), NaH2PO4 (pH 4.3), Na2HPO4 (pH 8.6) are high osmotic solutions. Used as. ACs could not be made with KNO3 (acidic), alkaline and NaCl solutions, but ACs appeared with acidic solutions of KH2PO4 and NaH2PO4 (pH 4.3). Thus, KNO3, which is acidic but does not contain PO43-, could not produce AC, suggesting that PO43- is more important than K + for the formation of AC.
In this regard, anionic drugs are believed to act primarily on the outer membrane surface of the erythrocyte membrane bilayer and expand it. However, the protrusions examined at this point had the same Hb concentration as the ACs body, and it was considered that the entire membrane containing Hb formed protrusions (Figure 1b and 1c). Immersing normal red blood cells in hyperosmotic fluid allows water to escape and form a sharp, pointed shape. On the other hand, red blood cells soaked in hypotonic fluid increase in volume and loosen the structure of the red blood cell membrane. When erythrocytes are exposed to high osmotic pressure in this state, the transmembrane proteins scattered in the erythrocyte membrane are positively charged by the acidity of the high osmotic solution. As a result, it was considered that a part of the transmembrane protein (+) was pulled out by the phosphate anion (PO43-) eluted from the erythrocytes, and a round protrusion was formed at the tip.
The anatomical changes in osmotic pressure before and after the hemolytic environment are similar. In the nephron simulation using urine, ACs were not observed until the red blood cells reached the hemolytic environment. This is true even after a hemolytic environment. Therefore, it is considered that the erythrocytes formed by bleeding from the glomerulus and renal tubules before reaching the hemolytic environment are degenerative, but the morphology formed by bleeding from the renal tubules after the hemolytic environment is homogeneous. Be done. Red blood cells formed by bleeding from the glomeruli and renal tubules prior to the hemolytic environment are considered to be degenerative, and the morphology formed by bleeding after the hemolytic environment, including the urinary tract, is considered to be uniform.
5. Regarding erythrocyte morphology:
Typical erythrocyte morphologies of deformity include spiny, rinng, and target erythrocytes. Of these, the spiny type is the most characteristic and specific form, and is closely associated with glomerular disease, Köhler 20) states. On the other hand, in non-glomerular hematuria, “ACs-like cells” appear when red blood cells are immersed in the following single urine conditions. Urine: 745 mOsm (pH 5.0, 15 minutes), 273 mOsm (pH 5.0, 2 hours), 273 mOsm (pH 5.0, 2 hours), 385 mOsm (pH 5.5, 2 hours), 607 mOsm (pH 5.5, 2 hours), 607 mOsm (pH 5.0, 2 hours), 607 mOsm (pH 5.0, 2 hours) 5.5, 2 hours), 933 mOsm (pH 5.0, 2 hours), 312 mOsm (pH 5.0, room temperature overnight). These conditions can result in the development of “ACs-like cells” when ureteral stones or hematuria is stagnant in the bladder, and these red blood cells should be classified as uniform red blood cells.
Conclusion
From the above results, the author proposes the following mechanism of his ACs formation in vivo.
Red blood cells due to bleeding from the renal tubules before reaching the glomerulus and hemolytic environment reach the hemolytic environment. Here, erythrocytes become erythrocytes with various Hb concentrations and contract when the hemolyzed erythrocytes reach the hyperosmolar region. This contraction acts disproportionately depending on the degree of damage to the hemolytic erythrocyte membrane, leading to the formation of dysmorphic erythrocytes such as ACs, ring-shaped cells, and target cells.
Acknowledgments
We would like to thank Dr. Hiroshi Sato of Tohoku University for lending us a valuable list of renal biopsies to prepare our paper.
References
- Birch DF, Fairley KF, Haematuria: Glomerular or non- glomerular? Lancet 1979; 314(6147): 845-6.
- Schramek P, Moritsch A, Haschkowitz H. In vitro generation of dysmorphic erythrocytes. Kidney Int 1989; 36: 72-7.PubMed
- Lin JT, Wada H, Maeda H, et al. Mechanism of Hematuria in Glomerular Disease. An electron microscopic study in a case of diffuse membranous glomerulonephritis. Nephron 1983; 35(1): 68-72PubMed
- Makino H, Nishimura S, Takaoka M, et al. Mechanism of Hematuria. II. A scanning electron microscopic demonstration of the passage of blood cells through a glomerular capillary wall in rabbit Masugi nephritis. Nephron 1988; 50(2): 142-50.PubMed
- Stapleton FB. Morphology of urinary erythrocytes: A simple guide in localizing the site of hematuria. Pediatr Clin North Am 1987; 34(3): 561-9PubMed
- Kubota H, Yamabe H, Ozawa K, et al. Mechanism of urinary erythrocyte deformity in patients with glomerular disease.Nephron 1988; 48(4): 338-9.PubMed
- Daza JL, De Rosa M, De Rosa G. Dysmorphic red blood cell formation. Cleve Clin J Med. 2018; 85(1):12-13. doi: 10.3949/ccjm.85a.17029.PubMed
- Yuste C, Gutierrez E, Sevillano AM, et al. Pathogenesis of glomerular haematuria. World J Nephrol 2015; 4(2): 185-95. doi: 10.5527/wjn.v4.i2.185.PubMed
- Miura H, Yamaguchi I, Fukuyama H, et al. Causative mechanism of dysmorphic erythrocytes in urine. Japanese Journal of Medical Technology 1992; 41, 733-9.J-STAGE
- Miura H, Yamaguchi I, Mityzawa K. Causative mechanism of dysmorphic erythrocytess in urine. 2nd Report: Effect of various elements in low osmotic solutions. Japanese Journal of Medical Technology, 1993; 42, 1479-84.PubMed
- Miura H, Yamaguchi I, Fukuyama H, et al. Causative mechanism of dysmorphic erythrocytes in urine. 3rd report Effect of various elements in high osmotic solutions. Japanese Journal of Medical Technology, 1994; 43: 1735-41.
- Kitamoto Y, Yide C, Tomita M, et al. The mechanism of glomerular dysmorphic red cell formation in the kidney. Tohoku J. Exp. Med 1992; 167(2): 93-105.PubMed
- Rath B, Turner C, Hartley B, et al. What makes red cells dysmorphic in glomerular haematuria? Pediatr Nephrol 1992; 6(5): 424-7.
- Briner VA, Reinhart WH. In vitro production of glomerular red cells: role of pH and osmolality. Nephron 1990; 56(1): 13-8.PubMed
- Halbhuber KJ, Fünfstück R, Kühn B, et al. Dysmorphic red cell formation in glomerulonephritis 2. In vitro generation of erythrocytes. Cell Mol Biol 1994; 40(8): e1125-33.PubMed
- Palygin O, Ilatovskaya DV, Staruschenko A. Protease-activated receptors in kidney disease progression. Am J Physiol Renal Physiol 2016 Dec 1; 311(6): F1140-4. doi: 10.1152/ajprenal.00460.2016. Epub 2016 Oct 12.PubMed
- Degrell P, Wagner Z, Szijarto IA, et al. Morphology of glomerular hematuria is reproduced in vitro by carbonyl stress. Nephron Exp Nephrol 2008; 110(1): e25-30. doi: 10.1159/000151437. Epub 2008 Aug 18.PubMed
- Kitamoto Y, Arizono K, Fukui H, et al. Urinary thrombin: A novel marker of glomerular inflammation for the diagnosis of crescentic glomerulonephritis (prospective observational study). PLoS One 2015; 10(3): e0118704.PubMed
- Birch DF, Fairley KF, Becker GJ, et al. A color atlas of urine microscopy. P27. (Chapman and Hall medical Tokyo 1994)
- Köhler H, Wandel E, Brunck B. Acanthocyturia- A characterristic marker for glomerular bleeding. Kidney Int 1991; 40(1): 115-20PubMed


